葡萄糖是所有生物异养生长最重要的碳和能量来源之一。然而,据报道,葡萄糖是支持热球菌目超嗜热古菌生长的不良底物。为了促进嗜热球菌NA1的葡萄糖辅助生长,采用了适应性进化过程。在工业应用的努力中,葡萄糖适应细胞进一步测试了生产氢气的潜力,利用食物加工废物作为含有葡萄糖组成的多糖的有前途的零值底物。
将T. onnurineus NA1细胞转移到含葡萄糖的新鲜培养基中,直到细胞生长增加,进行适应性进化。进行基因组测序以确定适应细胞的遗传变化。以葡萄糖或马铃薯皮废料为底物,分析了亲本菌株和葡萄糖适应细胞的产氢量。
与亲本菌株相比,葡萄糖适应细胞WG-100T的细胞密度和葡萄糖消耗分别增加了10.8倍和14.7倍。WG-100T的基因组测序显示,共有17个基因发生了变化,包括编码转录因子的基因和参与各种运输系统的几种蛋白质。WG-100T利用马铃薯皮废弃物同时糖化发酵制氢。
本研究表明,热球菌菌株的性能通过适应性进化得到改善,导致更快地利用葡萄糖。此外,研究表明,使用超嗜热菌可以在不预处理食品加工废弃物进行糖化的情况下产生生物氢。
一种超嗜热古菌通过适应性进化来提高葡萄糖的利用,证明了在没有糖化预处理的情况下从马铃薯皮废物中产生氢气的潜力。这一策略被认为有助于扩大利用各种生物质废弃物生产增值产品的糖基细胞工厂的范围。
由于气候变化和能源危机,氢气(H2)引起了人们的关注。目前,96%的氢气来自化石燃料,因此需要利用可再生资源生产氢气的技术。利用食品加工过程中产生的农产品残留物或废弃物进行暗发酵生产生物氢被认为是一种替代方法[1,2]。马铃薯皮废料每年在工业过程中产生7 - 14万吨,含有大量淀粉和非淀粉多糖等碳水化合物以及蛋白质和木质素等其他成分[3,4]。因此,它被认为是生产各种增值产品(如生物燃料、抗氧化剂和生物吸附剂)的有前途的零价值底物[5,6,7,8,9]。然而,为了让大多数微生物利用马铃薯皮,糖化需要酸或酶水解、热解或水热处理等预处理工艺。另一种方法可能是使用嗜热菌或超嗜热菌与嗜热酶在高温下降解多糖,其中糖化和发酵可以通过底物液化同时发生。
葡萄糖是淀粉的关键成分,是支持所有生物异养生长的重要碳和能量来源之一[10]。以葡萄糖为底物的超嗜热菌的生长研究主要报道在Thermotoga和Sulfolobus两种中[11,12,13,14]。然而,据报道,与麦芽糖和淀粉等其他糖相比,在葡萄糖[15]存在的情况下,海洋热藓的生长速度较低[16]。关于葡萄糖辅助其他超嗜热菌生长的研究非常有限,有报道称furiosus焦球菌(Pyrococcus furiosus)和Aeropyrum pernix对葡萄糖生长缓慢[17,18]。onnurineus热球菌NA1属于热球菌目,已知利用淀粉、甲酸盐和一氧化碳(CO)等底物产生生物氢[19,20,21]。许多研究都试图通过适应性实验室进化和基因工程来提高该物种的性能,并显著提高了其生长和H2产量[22,23,24,25]。然而,尽管它的胃口多种多样,但对葡萄糖的生长却从未确定过。
在这项研究中,我们研究了T. onnurineus NA1在葡萄糖上生长的潜力,并利用一个适应性进化过程来增强葡萄糖辅助生长。我们进一步测试了利用马铃薯皮废物在葡萄糖适应细胞中产生氢气。
从深海热液喷口区分离的onnurineus热球菌NA1[26]在改良培养基1 (MM1)中作为基础培养基[20],在80°C, pH 6.5的条件下常规培养。为了在血清瓶中培养,将葡萄糖原液添加到最终浓度为2、5、10、15或20 g/L的葡萄糖,在含有缺氧气体混合物(N2:H2=95:5)的厌氧室(Coy实验室产品,Grass Lake,美国)中制备MM1 +葡萄糖培养基。将200 g/L葡萄糖溶于水中,蒸压制备葡萄糖原液。分别加入5 g/L麦芽糖、淀粉或麦芽糖糊精配制MM1 +碳水化合物培养基。加入终浓度为0.005%的Na2S?9H2O,去除培养基中的溶解氧。
烹饪前产生的土豆皮废料由当地一家餐馆提供。干燥马铃薯皮(DPP)是将马铃薯皮在60℃下干燥24 h,在实验室研磨后制成,在- 20℃保存后使用。血清瓶分批培养时,将DPP加入血清瓶终浓度为3.3、5.0或6.6 g/L的MM1 + DPP培养基中,用2n HCl调节pH至6.5。
如前所述[25],在生物反应器中进行批量培养。分别加入20 g/L葡萄糖和3.3 g/L DPP制备MM1 +葡萄糖和MM1 + DPP培养基,用纯度为99.999%的纯氩气在80℃下净化60 min,不进行高压灭菌。用5n NaOH控制pH在6.1 ~ 6.2之间。
在含有20 g/L葡萄糖的MM1 +葡萄糖培养基中进行适应进化。亲本菌株在血清瓶中80℃培养24 h后,用无菌注射器将2% (v/v)的培养液转移到新鲜培养基中。此转移步骤重复100次。
利用BLAST (Basic Local Alignment search Tool)对美国国家生物技术信息中心(NCBI, Bethesda, USA)的非冗余蛋白数据库进行氨基酸序列的同源性检索。使用EMBL-EBI (https://www.ebi.ac.uk)提供的T-Coffee程序进行多序列比对。采用苯酚/氯仿/异戊醇(25:24:1,v/v)提取和纯化基因组DNA。基因组测序采用Pacific Biosciences (Menlo Park, USA)的单分子实时(SMRT)测序技术,采用10kb插入文库,覆盖范围约为100倍[27]。如先前报道的那样进行装配和一致抛光[28]。所有突变通过PCR和Sanger测序进行验证,引物见补充材料表S1。
利用紫外-可见分光光度计(BioPhotometer plus;埃彭多夫,汉堡,德国)。葡萄糖和乙酸的浓度采用hplc -折射率检测器(YL 9100;YL仪器公司,安阳,大韩民国)配备Rezex ROA色谱柱(300 × 7.8 mm;Phenomenex,托伦斯,美国)。HPLC的流动相为2.5 mM硫酸,流速为0.6 mL/min。H2浓度采用YL6100气相色谱仪(YL Instrument Co., Anyang, Republic of Korea),配备Molsieve 5a和Porapak N色谱柱(Supelco, Bellefonte, USA),导热系数和火焰电离检测器。以氩气为载气,流速为30ml /min。使用湿式气计(品川,东京,日本)测量生物反应器出口气体的总量。
摘要
新颖性声明
介绍
材料与方法
结果与讨论
结论
数据可用性
参考文献
作者信息
道德声明
补充信息
搜索
导航
#####
研究了嗜热古细菌T. onnurineus NA1是否能以葡萄糖为底物生长。随着葡萄糖浓度从2 g/L增加到20 g/L,细胞密度逐渐增加(图1a)。葡萄糖消耗实际上在时间过程曲线中与细胞生长成正比(图1b)。然而,据报道,P. furiosus和A. pernix的细胞密度较低,仅消耗少量葡萄糖[17,18]。
图1
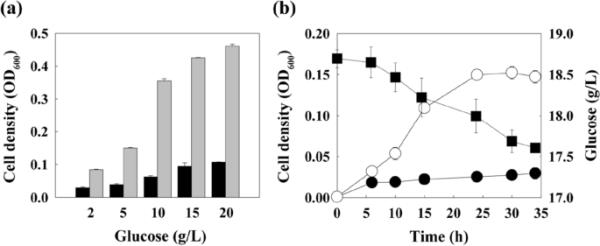
T. onnurineus NA1在葡萄糖作用下的细胞生长。a葡萄糖浓度对亲本菌株(黑色条)和葡萄糖适应的WG-100T细胞(灰色条)生长的影响。b亲本菌株在20 g/L葡萄糖(封闭圆圈)和剩余葡萄糖(封闭方块)存在或不存在时细胞生长的时间过程曲线。误差条表示两个(a)或三个(b)生物重复的标准偏差。OD600, 600 nm光密度
有报道称,T. maritima通过适应性进化提高了生长速度和葡萄糖利用[29]。因此,该方法适用于T. onnurineus NA1。培养24小时后,将细胞转移到含有20 g/L葡萄糖的新鲜培养基中,以适应葡萄糖。随着连续转移的进行,观察到细胞密度和葡萄糖消耗逐渐增加(数据未显示)。第100个转移的细胞,命名为WG-100T,在所有测试的葡萄糖浓度下,与亲本菌株的相应值相比,细胞密度高2.9-5.7倍(图1a)。
在pH维持在6.1-6.2的生物反应器中,对进化细胞的表型特性进行了进一步的表征。适应后的细胞表现出明显的生长改善,细胞密度增加了10.8倍(图2a),葡萄糖消耗增加了14.9倍(图2b)。与亲本菌株相比,适应细胞培养中乙酸酯(图2c)和H2(图2d)的代谢物产量分别增加了15.7倍和14.2倍。亲本菌株每消耗葡萄糖产生的乙酸和H2的摩尔比为1.5和1.8,适应细胞为1.6和1.7。WG-100T在特定生长率、最大葡萄糖消耗率、最大乙酸产量和最大H2产量上分别比亲本菌株提高2.2倍、8.5倍、15.2倍和15.6倍(表1)。在生物量、乙酸和H2产量方面,WG-100T分别比亲本菌株提高22.3倍、3.3倍和5.8倍。WG-100T在简单分批培养中获得的生物量生产力为0.16 g/L/h,与分批补料培养Saccharolobus solfataricus P2或Saccharolobus solfataricus Gθ获得的高细胞密度相当[30,31]。
图2
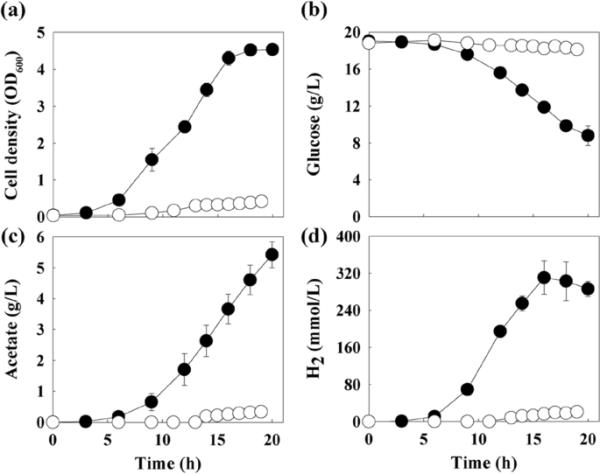
葡萄糖适应WG-100T细胞在生物反应器中的批量培养概况。测定亲本菌株(开圈)和WG-100T(闭圈)的细胞密度(a)、剩余葡萄糖浓度(b)、乙酸浓度(c)和H2浓度(d)。误差条表示三个独立培养的标准差。OD600, 600 nm光密度
表1母菌株与WG-100T的动力学参数比较
为了了解适应细胞如何在葡萄糖存在下获得增强的生长表型,通过基因组测序分析了基因组中的遗传变异。在WG-100T基因组中共检测到17个突变(表2)。尽管大多数突变与甲酸或一氧化碳条件下进化的细胞中发现的突变不同[22,23],编码芳香氨基酸渗透酶的TON_0820基因的点突变在所有三种条件下都被检测到。有趣的是,突变位置都不同[22,23]:基因在葡萄糖条件下的适应性进化过程中携带了一个移框突变。由于该突变已被证实在CO条件下刺激生长,因此它可能对葡萄糖条件下的细胞生长产生积极或消极的影响。发现几个突变基因与转运系统有关:abc型转运体渗透酶(TON_0597)、质子/谷氨酸同向转运体(TON_0901)、abc型麦芽糊精转运系统(TON_1795)和黄嘌呤/尿嘧啶渗透酶(TON_1957)。TON_1795和TON_1957有帧移突变,可能导致无功能基因产物。编码渗透酶的TON_0597基因属于编码糖结合蛋白(TON_0594)、atp酶(TON_0595)、渗透酶(TON_0596)的基因簇,是atp结合盒(ABC)型转运体系统的组成部分。TON_0594的底物特异性尚不清楚,但似乎与糖有关。TON_0597基因的突变可能会增加对生长所需底物的吸收,从而导致WG-100T的生长改善。TON_0901与kodakarensis热球菌和horikoshii热球菌的钠偶联天冬氨酸转运体分别具有78.3%和70.6%的氨基酸序列同源性,已被功能和结构鉴定为特异性运输天冬氨酸[32,33,34]。所有与底物天冬氨酸相互作用的残基都是严格保守的(图S1),因此TON_0901的突变可能通过增加天冬氨酸的摄取来促进细胞生长。为了确定TON_0597或TON_0901位点的改变对所观察到的适应细胞表型的影响,构建具有每个突变的突变体是有帮助的。
表2 WG-100T基因组变化总结
此外,需要多组学分析来确定解释进化的T. onnurineus NA1细胞表型的机制。各种基因表达水平的变化,包括那些编码转运蛋白的基因,可能起着有益的作用。这可能有助于鉴定T. onnurineus NA1中的葡萄糖转运基因。已经通过基因表达分析预测了酸硫藻(Sulfolobus acidocalarius)假定的葡萄糖ABC转运基因簇,因为它在葡萄糖存在下的生长过程中表现出上调[35]。WG-100T表型变化的机制有待进一步研究。
基于T. onnurineus NA1通过适应性进化增强了对葡萄糖的生长,研究了WG-100T对葡萄糖、麦芽糖、麦芽糖糊精和淀粉组成的多种糖类的生长能力。与亲本菌株相比,在存在麦芽糖的情况下,WG-100T的生长没有增强(图3)。另一方面,在存在淀粉和麦芽糖糊精的情况下,WG-100T的细胞密度分别比亲本菌株高1.2倍和2.3倍(图3a)。氢气产量随细胞密度的增加成比例增加(图3b)。这些结果表明,糖适应细胞比亲本菌株更有效地利用低聚糖生产H2。
图3
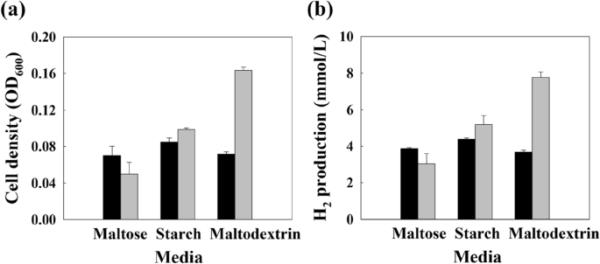
碳水化合物作为底物存在下WG-100T细胞的生长。麦芽糖、淀粉和麦芽糖糊精对亲本菌株(黑条)和WG-100T(灰条)细胞生长(a)和产氢(b)的影响。误差条表示两个生物重复的标准差。OD600, 600 nm光密度
以马铃薯皮渣为底物,利用T. onnurineus NA1亲本菌株和WG-100T产氢试验。将马铃薯皮晒干并磨碎后加入血清瓶中,使亲本菌株和WG-100T产生H2。随着干马铃薯皮(DPP)含量从3.3 g/L增加到6.7 g/L,两种细胞的H2产率均下降。据报道,马铃薯皮中的有毒化合物可能会影响微生物的生长[36,37]。同时,WG-100T的产氢率比亲本菌株高1.3 ~ 1.5倍(图4a)。为了克服pH下降造成的生长限制,我们尝试在pH-stat生物反应器中进行培养,结果发现与血清瓶相比,H2产率有所提高(图4b)。
图4

利用马铃薯皮废渣生产WG-100T制氢。a在含有不同量马铃薯皮废料的血清瓶中,亲本菌株(开圈)和WG-100T(闭圈)的H2产量。b亲本菌株(开圈)和WG-100T(闭圈)在含3.3 g/L马铃薯皮废弃物的pH-stat生物反应器中产氢情况。误差条表示两个生物重复的标准差
WG-100T在产氢率(最高为2.83 vs. 0.97 mmol/L/h)和产氢率(1.29 vs. 0.37 mmol/L/h)方面均显著优于亲本菌株(表1)。结果表明,对葡萄糖的适应增强了菌株对含碳水化合物的食物垃圾的消化能力。值得注意的是,超嗜热古细菌T. onnurineus NA1利用经过低温干燥和研磨预处理的马铃薯皮废料制取H2。与利用马铃薯加工业产生的马铃薯蒸汽皮的caldiclulosiruptor saccharolyticus和Thermotoga neapolitana产氢速率相比[7],WG-100T的单位底物产氢速率降低了2倍。目前,很难说这种差异是由于马铃薯皮的预处理还是菌株的能力。
嗜热球菌NA1被证实能够在葡萄糖上生长,但其对葡萄糖的性能不如利用葡萄糖的嗜热细菌。通过适应性进化,微生物找到了更有效地利用葡萄糖的方法。基因组测序揭示了适应细胞的基因组变化。进一步的研究是必要的,以确定分子的作用机制,可以解释适应细胞的生理变化。通过同时糖化和发酵生产H2,确定了超嗜热古菌作为马铃薯皮废弃物的潜在利用菌。基因工程的进一步应用将有助于提高氢气产量。
以下是电子补充材料的链接。
下载原文档:https://link.springer.com/content/pdf/10.1007/s12649-023-02049-z.pdf
为您推荐:
- 【老鹰对雄鹿的大小比分预测,老鹰对雄鹿比赛结果】 2025-12-17
- 韩国网红年入破亿成常态!揭秘YouTube暴富潮背后的惊人真相?韩国一个网红 2025-12-17
- 【电脑游戏排名,电脑游戏排名榜2025前十名手游】 2025-12-17
- 火箭vs灰熊?火箭VS灰熊全场回放 2025-12-17
- 【春秋霸业,春秋霸业一壶茶的意思】 2025-12-17
- 湖人半场81分?湖人半场领先33分新闻 2025-12-17


